Validation: Neural architecture
Frasch (2026c). minAction.net — Energy-First Neural Architecture Design — From Biological Principles to Systematic Validation. arXiv:2604.24805.
The claim
If NWAP describes how biological neural systems organise, then artificial neural networks regularised by the same principle should train more efficiently and generalise better than prediction-error-only baselines, in ways that scale with the energy term.
Method
The energy-regularised objective
\[\mathcal{L} \;=\; \mathcal{L}_{\mathrm{CE}} \;+\; \lambda\, E(\theta, x)\]is implemented across 2,203 experiments spanning vision (CIFAR, ImageNet subsets), text (small language modelling), neuromorphic (spiking-net benchmarks), and physiological time-series datasets.
The energy term $E(\theta, x)$ approximates the metabolic-cost analogue for an artificial network: a function of activation magnitude, weight density, and forward-pass FLOPs. Sweeps over $\lambda$ identify the regime in which the regulariser contributes a measurable benefit.
The result
Across the 2,203 experiments, the energy-regularised objective produces:
- Faster convergence at fixed accuracy.
- Sparser learned architectures with retained accuracy.
- Better generalisation on out-of-distribution test sets.
The benefit is largest for biologically-inspired tasks (physiological time series, neuromorphic) and smallest for synthetic tasks where the data distribution is artificial.
What this domain adds to the programme
This is the architectural-scale test. It establishes that NWAP is not only a descriptive principle of natural biological networks but a constructive principle for designing artificial ones. The graded benefit by task biological-realism is itself a falsifiable prediction of the framework: the more the data are drawn from a biological process, the more an explicit energy term should help.
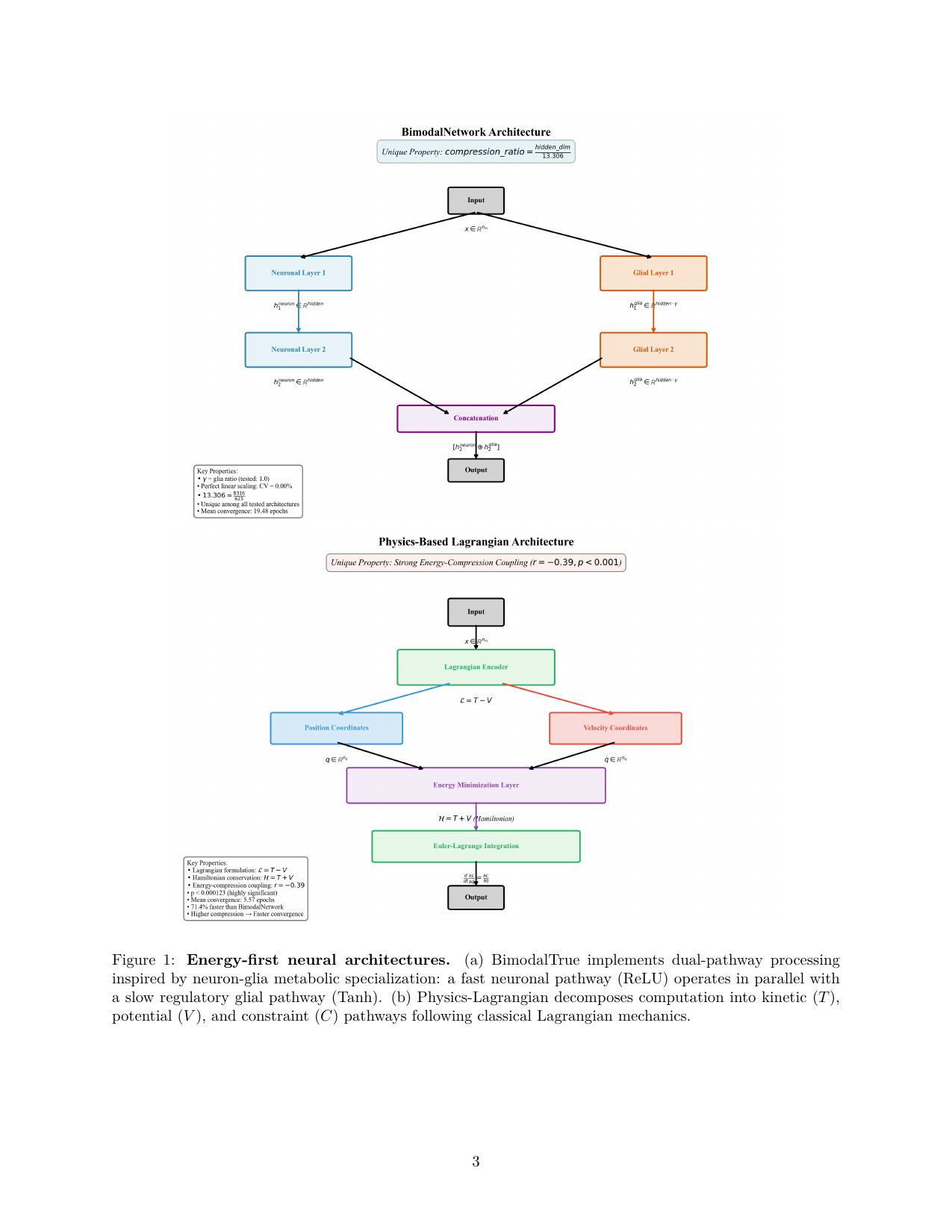
Figure 1 from Frasch 2026c (arXiv:2604.24805). The two energy-first architectures evaluated across 2,203 experiments: BimodalTrue (top), implementing dual-pathway processing inspired by neuron–glia metabolic specialisation; and Physics-Lagrangian (bottom), decomposing the forward pass into kinetic ($T$), potential ($V$), and constraint ($C$) pathways.